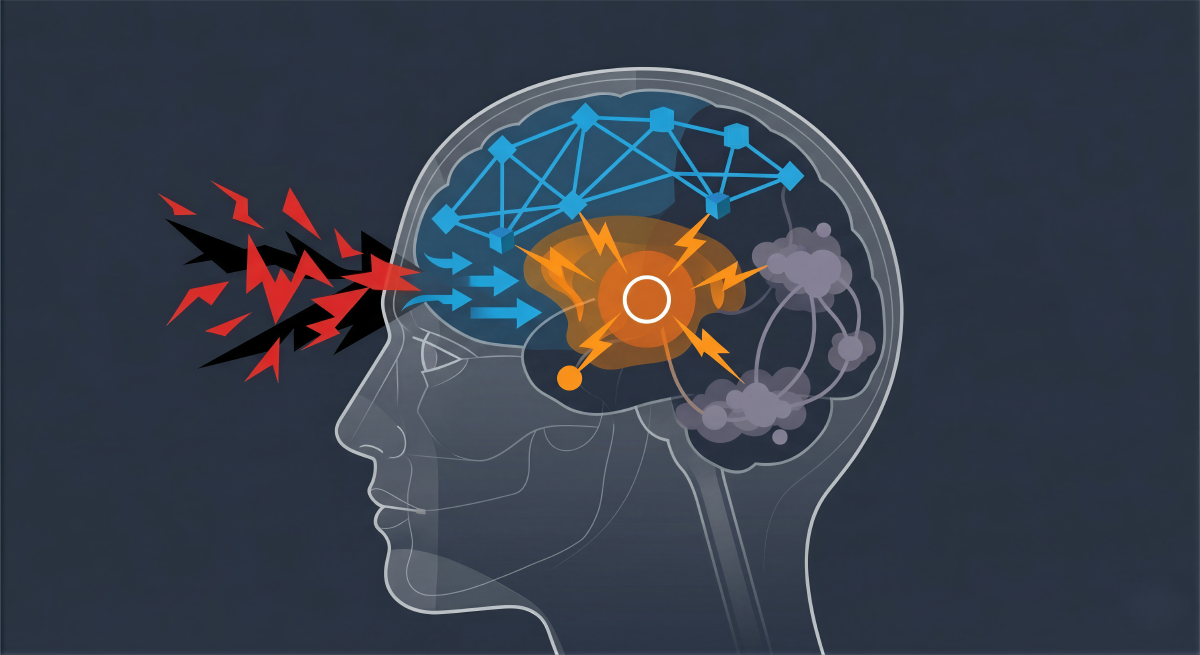
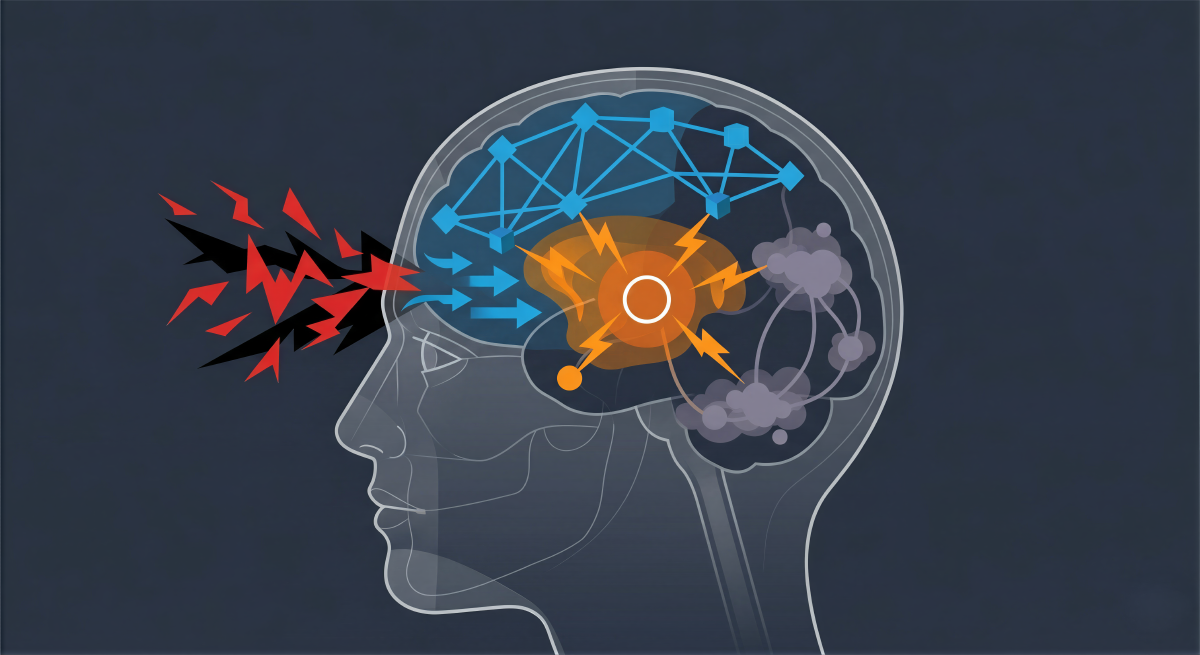
Introduction: The Neurobiological Crucible of Crisis#
In an era defined by extreme volatility, uncertainty, complexity, and ambiguity, the probability of systemic crises unfolding continues to accelerate. When global disruptions, catastrophic infrastructure failures, or acute market shocks materialize, the most immediate and profound structural collapse often occurs not in the physical or economic environment but in the human cognitive architecture. The modern crisis environment demands that leaders process highly ambiguous information, rapidly discard obsolete mental models, and execute high-velocity, high-stakes decisions. Yet, the evolutionary design of the human brain actively opposes this requirement. When confronted with an overwhelming threat or a profoundly unpredictable environment, the brain’s instinctual, neurobiological response is to bypass the metabolically expensive, computationally heavy networks responsible for fluid cognitive agility. Instead, it reverts to rigid, historically ingrained habits, manifesting behaviorally as either paralyzing panic or a maladaptive hyper-focus on irrelevant details.
The capacity to resist this neurochemical hijacking and to maintain goal-directed, adaptable decision-making when predictive models fail is fundamentally rooted in a definable, observable, and trainable neurobiological mechanism known as cognitive flexibility, or set-shifting. Cognitive flexibility is the executive capacity to switch between different modes of thinking, shift perspectives in response to changing environmental demands, and find workable solutions to conflicting problems. It is the neurological antidote to stress-induced rigidity.
This exhaustive research report delivers a deep and nuanced examination of the neuro-mechanics of crisis leadership. By deconstructing the brain’s molecular responses to acute stress, mapping the architecture of intrinsic executive-function networks, exploring the Bayesian mechanics of predictive coding, and examining historical paradigms of extreme crisis management, this analysis outlines the precise mechanisms underlying the agile mind. Furthermore, it provides a comprehensive framework detailing how master architects of leadership explicitly and implicitly train their prefrontal networks to maintain supreme cognitive stability in the eye of the storm.
The Neurochemical Architecture of Acute Stress#
To fundamentally understand cognitive agility, it is first necessary to dissect the neurochemical siege that occurs during acute stress. The human brain operates as a hierarchical prediction engine, and the undisputed apex of this hierarchy is the prefrontal cortex (PFC). Subserving the highest-order cognitive abilities, the PFC is responsible for working memory, goal-directed behavior, impulse control, emotional regulation, and the dynamic allocation of attention. However, this advanced computational capacity comes at a severe biological cost: the PFC is uniquely fragile. Its intricate network connections are tremendously dependent on a highly specific and tightly regulated neurochemical environment, making it the brain region most acutely sensitive to the detrimental effects of stress exposure.
Catecholaminergic Surges and Prefrontal Decoupling#
The functional integrity of the PFC is regulated by ascending arousal pathways that release catecholamines, specifically norepinephrine (NE) and dopamine (DA), into the cortical environment. Under optimal, non-threatening conditions, such as states of alertness or engaged interest, moderate levels of these neurotransmitters are released. These molecules engage postsynaptic receptors (such as alpha-2A receptors located on the dendritic spines of PFC pyramidal cells), effectively reinforcing the synaptic network connectivity that sustains working memory and top-down executive control. This dynamic represents the apex of the inverted-U curve of arousal, the optimal state where cognitive performance, strategic planning, and leadership efficacy are maximized.
However, the onset of an acute, uncontrollable crisis fundamentally alters this delicate neurochemical milieu. When a leader perceives a situation as profoundly threatening and, critically, out of their immediate control, the brainstem floods the cortex with exceptionally high levels of catecholamines. This massive molecular surge rapidly and violently switches the brain’s operational state from a thoughtful, reflective, and analytical mode to an unconscious, reflexive, and reactive state. High levels of D1 receptor stimulation and excessive norepinephrine release weaken synaptic connections in the prefrontal cortex, effectively taking the executive center “off-line” and allowing more primitive subcortical structures to assume command.
This rapid prefrontal decoupling is simultaneously accompanied and amplified by an influx of glucocorticoids, such as cortisol, which coordinate and exaggerate the switch by binding to receptors in both the PFC and the subcortical amygdala. The consequence is a rapid architectural reorganization of cognitive resources. The PFC loses its top-down regulatory control, meaning the individual loses access to complex problem-solving capabilities and nuanced working memory. Simultaneously, affective processing in the amygdala (the brain’s fear center) and habitual, reflexive responses in the basal ganglia (specifically the striatum) are dramatically strengthened and sensitized.
The Functional Shift from Goal-Directed to Habitual Systems#
This neurochemical reorganization orchestrates a profound transition in the mechanics of human decision-making. Advanced decision-making frameworks operate on a spectrum between two distinct computational systems: goal-directed (model-based) processing and habitual (model-free) processing.
Goal-directed behavior is highly flexible. It relies on forming a continuously updated internal model of the environment, evaluating the future consequences of potential actions, and rapidly adapting to changing rules or contingencies. The prefrontal cortex and the dorsomedial striatum predominantly mediate it. Conversely, habitual behavior relies on stimulus-response associations that have been deeply reinforced over historical time. Habitual processing is computationally efficient and metabolically inexpensive, but it is highly rigid, deeply perseverative, and wholly insensitive to sudden changes in outcome valuation. This system is anchored in the dorsolateral striatum (the putamen) and the amygdala.
Under acute stress, the structural impairment of the PFC forces the cognitive apparatus to default entirely to the habitual system. Clinical studies utilizing acute psychosocial stress paradigms, such as the Socially Evaluated Cold Pressor Task (SECPT), consistently demonstrate that physiological stress impairs the flexible implementation of task goals, disrupts context-dependent memory, and induces a prominent, measurable shift toward habitual, model-free strategies. In experimental setups such as the Two-Step Markov Task, stressed individuals show a marked decrease in model-based learning and instead rely on rigid, previously rewarded actions regardless of new environmental constraints.
For a leader navigating a systemic crisis, this neurobiological shift is often the catalyst for operational catastrophe. Crises, by their very definition, present novel, unprecedented environmental configurations where historical heuristics, standard operating procedures, and familiar playbooks are explicitly invalid. When the stressed brain defaults to habit, it executes obsolete responses. This biological reality explains the phenomena of “threat-rigidity” in corporate leadership, resulting in a rigid hyper-focus on irrelevant, familiar tasks, an inability to process disconfirming evidence, or a panic-driven paralysis.
To understand the neurobiological shift that occurs during a crisis, it is essential to contrast the brain’s functioning between two distinct operational modes: the optimal, goal-directed system and the stress-induced, habit-based system.
- Primary Neural Correlates: In a pre-crisis state, the goal-directed system relies heavily on the prefrontal cortex, dorsomedial striatum, and hippocampus. Under acute stress, control shifts to the habit-based system, driven by the amygdala and dorsolateral striatum (putamen).
- Computational Model: The optimal brain operates on a model-based system that continuously evaluates future states and probabilistic outcomes. Conversely, the stressed brain defaults to a model-free system, relying on historically reinforced stimulus-response reactions.
- Cognitive Flexibility: Goal-directed processing maintains high flexibility, allowing rapid adaptation to new rules, constraints, and data. Habit-based processing is marked by low flexibility, resulting in perseveration and a profound resistance to shifting paradigms.
- Metabolic Cost: The goal-directed system demands a high metabolic cost to sustain active working memory and focused attention. The habit-based system is metabolically cheap, conserving energy through automatic, reflexive execution.
- Leadership Manifestation: While optimal functioning enables strategic pivoting, creative problem-solving, and sharp situational awareness, a stress-induced habit state typically manifests as threat-rigidity, micromanagement, and a strict reliance on obsolete playbooks.
Crucially, the structural and functional changes triggered by acute stress are largely reversible, and decision-making can return to a goal-directed state once the stressor abates and the neurochemical balance is restored. The hallmark of extraordinary crisis leadership, therefore, lies not in the complete absence of the human stress response but in the trained neurological capacity to override this biological default. The agile mind preserves PFC function and goal-directed processing precisely when the catecholaminergic surge is attempting to shut it down.
The Architecture of Cognitive Flexibility: Networks of the Agile Mind#
Maintaining goal-directed control under pressure requires the continuous, uninterrupted operation of cognitive flexibility, a capacity operationalized in cognitive neuroscience as “set-shifting.” Cognitive flexibility is the higher-order executive ability to shift attention between multiple tasks, operations, or mental sets in response to shifting environmental demands. It requires discarding an obsolete perspective and rapidly adopting a new one to reduce cognitive conflict.
To fully grasp how master architects of leadership maneuver through high-stakes crises, it is imperative to analyze the large-scale intrinsic connectivity networks that facilitate set-shifting. Cognitive agility is not localized to a single cortical locus; rather, it emerges dynamically from the complex interplay, coupling, and decoupling of three primary neurocognitive networks: the Default Mode Network (DMN), the Central Executive Network (CEN) or Lateral Frontoparietal Network (L-FPN), and the Salience Network (SN).
The Central Executive and Default Mode Networks#
The Central Executive Network (CEN), which heavily overlaps with the Lateral Frontoparietal Network (L-FPN), is the neurological engine of active problem-solving and cognitive control. Anchored by the dorsolateral prefrontal cortex (dlPFC), the ventrolateral prefrontal cortex, and the posterior parietal cortex, this network is robustly engaged during cognitively demanding tasks requiring sustained attention, working memory, and rule-based decision-making. When a leader is actively “working the problem,” analyzing data, and formulating a strategic response, the CEN is highly activated.
Operating in direct opposition to the CEN is the Default Mode Network (DMN), a “task-negative” network anchored in the medial prefrontal cortex (mPFC) and the posterior cingulate cortex. The DMN is primarily active during rest, mind-wandering, internal reflection, autobiographical memory retrieval, and unconstrained rumination. For highly efficient, externally focused cognition to occur, the CEN and DMN must be strictly anti-correlated. When an individual engages in active problem-solving in a crisis, the CEN must fully activate, and the DMN must correspondingly deactivate to prevent internal distraction and anxious rumination.
The Orchestrating Role of the Salience Network#
The seamless, high-velocity transition between internal reflection (DMN) and active, external problem-solving (CEN) is orchestrated by the Salience Network (SN), frequently referred to in the literature as the Midcingulo-insular network (M-CIN). The SN serves as the brain’s central switchboard, anchored by the dorsal anterior cingulate cortex (dACC) and the anterior insula, and exhibits robust connectivity with subcortical and limbic structures, including the amygdala.
The primary function of the Salience Network is to continually monitor internal homeostatic states and external sensory inputs, identifying which stimuli in the environment are most “salient,” critical, or threatening. When a crisis erupts, the SN detects the profound anomaly, generates an immediate arousal signal, and performs a dual operation: it suppresses the internally focused Default Mode Network while simultaneously recruiting and activating the Central Executive Network to manage and resolve the threat.
Set-shifting, the biological core of cognitive agility, is highly dependent on the rapid, efficient, and conflict-free functionality of the dACC and its connectivity with the dlPFC. Complex set-shifting tasks require broad perspective changes and strong top-down biasing from the prefrontal cortex to resolve conflicting information streams. When a leader faces a rapidly evolving crisis, such as a catastrophic equipment failure, an unprecedented cyber-breach, or a sudden market collapse, the Salience Network must rapidly detach cognitive attention from the original strategic plan (the former mental set) and forcefully shift resources to the L-FPN to construct a novel, highly adaptive response strategy.
Dysfunction or dysregulation within these three networks explains the vast majority of leadership failures under pressure. If the Salience Network becomes hyperactive, driven by overwhelming, uncontrollable amygdala responses to perceived threat, it can fail to engage the CEN properly. This results in a state in which the leader is acutely and painfully aware of the crisis yet remains cognitively paralyzed, unable to formulate a coherent strategy. Conversely, factors such as aging, chronic stress, or psychopathology can lead to decreased coupling within the Salience Network itself and reduced decoupling from the DMN. Studies examining aging populations reveal that these specific network connectivity changes directly correlate with a measurable decline in cognitive flexibility, as evidenced by poor performance on the Trail Making Test and Color Trails Test, which in turn leads to increased perseverative behavior. Masterful crisis leadership requires maintaining robust, conflict-adaptation-dependent functional connectivity between the dACC and the right dlPFC, ensuring that shifts in strategy are executed smoothly and without perseverative drag.
Predictive Coding: The Bayesian Brain and the Calculus of Uncertainty#
To deeply understand why a crisis triggers such violent neurochemical and network-level disruptions in human beings, the phenomenon of leadership under pressure must be viewed through the advanced lens of computational neuroscience, specifically, the framework of predictive coding, active inference, and the Bayesian brain hypothesis.
The Brain as an Inference Engine#
The historical view of human perception posited that the brain is largely a passive receiver of sensory information, constructing its reality from the bottom up based on what the eyes see and the ears hear. The predictive coding framework, rooted in Bayesian probability theory, entirely inverts this paradigm. It proposes that the brain is a proactive, continuously calculating inference engine that operates to minimize surprise by actively generating internal models, top-down predictions, or “priors”, about what it expects to encounter in the sensorium.
Sensory input from the external world travels bottom-up. When the top-down predictions generated by the cortex precisely match the bottom-up sensory data received from the environment, the brain experiences minimal “prediction error,” and the internal model of reality is validated as accurate. However, when there is a significant discrepancy between what the brain expects to happen and what the environment provides, a “prediction error” is generated.
Operating under the fundamental biological law of the free energy principle, the brain seeks at all costs to minimize surprise, uncertainty, and entropy. Upon encountering a prediction error, the biological system must resolve the discrepancy to restore homeostasis. It accomplishes this through one of two primary mechanisms:
- Perceptual Inference (Belief Updating): The brain revises its internal predictions (its Bayesian priors) to align with the newly acquired sensory evidence, effectively updating its mental model of reality.
- Active Inference: The organism takes physical action in the environment, actively altering the sensory input so that it aligns with the preexisting prediction and reduces uncertainty about hidden states.
Crises as Cascading Prediction Errors#
In the context of predictive processing, a crisis is defined not by the physical damage it causes but by an environment characterized by extreme volatility that generates massive, unresolvable prediction errors. When global disruptions hit, the standard operational models and Bayesian priors that leaders rely upon to predict market behaviors, supply chains, or physical safety suddenly fail. The environment becomes fundamentally uncertain.
For a leader possessing high cognitive flexibility, prediction errors serve as vital, highly informative learning signals. The flexible brain accurately gauges the volatility of the new environment, decreases the “precision weighting” (the level of absolute confidence) of its prior beliefs, and relies heavily on incoming sensory data to rapidly update its mental models. This is the very essence of agility: recognizing instantly that the old map no longer matches the territory, accepting the prediction error, and drawing a new map in real-time.
However, the stressed, anxious, or rigid mind processes uncertainty through a fundamentally flawed calculus. Under acute anxiety or previous traumatic stress, the brain often develops “hyper-precise threat priors”. The anxious individual chronically overestimates the likelihood of harm, becoming hypersensitive to ambiguous stimuli and treating all uncertainty as an existential threat. When faced with a massive prediction error in a crisis, rather than engaging in perceptual inference and updating their beliefs, a process that requires high cognitive flexibility and deep PFC engagement, the rigid brain reinforces its prior beliefs.
This context rigidity and inability to down-regulate error signals lead directly to maladaptive active inference. The overwhelmed leader may engage in behavioral avoidance, extreme micromanagement, or outright denial. These actions are subconscious policies that attempt to artificially reduce volatility by limiting the leader’s access to corrective, yet highly uncomfortable, evidence. In predictive processing terms, the stress-induced shift from goal-directed to habit-based decision making is the brain’s desperate attempt to retreat to highly practiced, low-uncertainty priors, even when those priors are objectively useless for solving the current anomaly.
Furthermore, this framework extends beyond individual cognition to “second-order active interpersonal inference”, how a leader models the mental states of their team. During a crisis, a leader must not only update their model of the physical event but must also recursively infer how their team perceives the crisis and the leader’s response to it. Leaders lacking cognitive flexibility fail at this second-order inference, projecting their own hyper-precise threat priors onto their organization, thereby stifling psychological safety and cascading rigidity throughout the corporate structure.
High-Velocity Adaptive Cognition: Paradigms of Crisis Leadership#
Theoretical neuroscience provides the intricate cellular and computational foundation for cognitive agility, but its operational reality is best observed in acute, high-stakes environments. Examining historical anomalies where catastrophic prediction errors were met with exceptional executive control illuminates the practical mechanics of the agile mind.
Sullenberger and the OODA Loop of Flight 1549#
On January 15, 2009, US Airways Flight 1549 struck a dense flock of Canada geese shortly after takeoff from LaGuardia Airport, resulting in a dual-engine failure at a dangerously low altitude. Captain Chesley “Sully” Sullenberger and First Officer Jeffrey Skiles had precisely 208 seconds from the moment of the bird strike to the ditching of the Airbus A320 in the Hudson River.
In predictive coding terms, a total loss of thrust at 2,800 feet over a densely populated metropolis represents a catastrophic prediction error. The standard aviation “prior” strongly dictates that airplanes land on designated runways. The air traffic control tower, operating under this rigid protocol and physically removed from the cockpit’s immediate sensory data, repeatedly attempted to direct Flight 1549 back to LaGuardia or to nearby Teterboro Airport in New Jersey.
If Sullenberger had succumbed to the massive catecholaminergic surge expected in such a terrifying scenario, his prefrontal cortex would have decoupled, immediately transferring control to the habit-based striatum. The habitual response, reinforced by thousands of hours of standard operating procedures, would have been an obsessive, perseverative attempt to reach an airport, leading to an inevitable stall and a catastrophic urban crash. Instead, Sullenberger demonstrated supreme cognitive flexibility.
He engaged in what researchers term Naturalistic Decision-Making (NDM) and Recognition-Primed Decision-Making (RPDM). RPDM posits that experts do not make decisions in crises by systematically comparing all possible options. This process is computationally too slow and demands too much working memory for extreme emergencies. Instead, they draw on deep experiential reservoirs to rapidly match situational cues to successful action schemas. However, this specific situation was entirely novel; there was no exact pre-existing schema for a low-altitude dual-engine failure over Manhattan.
Crucially, Sullenberger utilized the behavioral manifestation of the Salience Network’s switching capability: the intentional pause. Neurocognitive research indicates that the simple act of pausing, even for a mere 50 to 100 milliseconds, allows the brain to inhibit habitual responses and focus cortical resources on the most relevant information. Sullenberger paused, suppressed the overwhelming threat signals from his amygdala, and executed a rapid epistemic evaluation of his parameters (altitude, airspeed, glide ratio). He broke the rigid mental set of “landing at an airport” (the essence of set-shifting) and formulated a novel, goal-directed model: a controlled ditching on the Hudson River.
This process perfectly maps onto the military decision-making framework known as the OODA Loop, originally developed by U.S. Air Force Colonel John Boyd.
To illustrate how Captain Sullenberger’s real-time decision-making aligns with both cognitive neuroscience and predictive processing frameworks, we can deconstruct his response through the four phases of the OODA Loop:
- Observe: Driven by the Salience Network (SN) threat detection, this phase involved acknowledging the bird strike, engine rollback, and catastrophic thrust loss. In predictive-processing terms, the sensory input radically violated the expected generative priors, resulting in a massive prediction error.
- Orient: Facilitated by the transition from the Default Mode Network (DMN) to the Central Executive Network (CEN), Sullenberger paused to assess altitude and distance while actively suppressing panic. This represents updating the generative model, assessing the precision weighting of the error, and suppressing DMN noise.
- Decide: Anchored in the Lateral Frontoparietal Network (L-FPN) and the dorsolateral prefrontal cortex (dlPFC), this phase required rapid set-shifting. By abandoning the LaGuardia prior and choosing the Hudson River as the only viable solution, Sullenberger generated a novel prior within the goal-directed system to efficiently resolve the error.
- Act: Relying on motor execution and working memory maintenance, the crew executed the “aviate-navigate-communicate” protocol, deployed the APU, and called for the checklist. This serves as the behavioral manifestation of active inference, altering the environment to align reality with the new predictive model.
Sullenberger’s ability to maintain executive control, start the auxiliary power unit (APU), and take manual control of the aircraft within just 18 seconds of the strike exemplifies the absolute pinnacle of goal-directed cognitive stability under toxic stress. It highlights how an experienced, well-trained crew can minimize cognitive effort, mitigate the effects of stress on attention, and achieve rapid leaps between states of knowledge.
Gene Kranz and the Distributed Cognition of Apollo 13#
While Flight 1549 highlights individual, high-velocity set-shifting, the Apollo 13 lunar mission crisis of 1970 demonstrates how cognitive flexibility must be applied to distributed organizational leadership. When an oxygen tank exploded en route to the moon, Mission Control in Houston was plunged into a scenario of profound volatility, extreme uncertainty, and severe resource constraint.
Lead Flight Director Gene Kranz faced a cascading series of prediction errors that violated every known parameter of spaceflight: rising CO2 levels, failing power grids, and an unviable trajectory. Rather than succumbing to organizational threat rigidity, Kranz engineered a communicative and operational environment that maximized his team’s collective prefrontal capacity. His famous directive, “Let’s work the problem, people. Let’s not make things worse by guessing,” is a masterclass in organizational cognitive regulation.
By demanding that engineers exclusively “work the problem” and explicitly avoid “guessing,” Kranz actively suppressed the Default Mode Network’s speculative, anxiety-driven noise and focused the team’s Central Executive Networks solely on data-driven, goal-directed processing. Furthermore, Kranz effectively managed cognitive load by defining rigid boundary conditions. In the famous CO2 scrubber adaptation scene, Kranz did not solve the engineering problem himself; he bounded the problem parameters (time limits, available onboard materials, risk tolerance) and orchestrated the experts within that strict cognitive framework.
This represents the strategic application of heuristics to deliberately constrain the search space of a problem, preventing the analysis paralysis that inevitably occurs when the prefrontal cortex is overwhelmed by complex, unbounded variables. By projecting outward calm and providing a clear, single-minded vision, Kranz fostered an environment of emotional safety that mitigated the catecholamine surge among his remote teams. This allowed them to collaborate, brainstorm, and engage in the rapid set-shifting required to adapt the Lunar Module into a lifeboat. Apollo 13 demonstrates that in massive organizational crises, leadership cognitive flexibility must scale; the leader must act as the ultimate Salience Network for the entire organization, identifying the critical objectives and dynamically shifting the collective attention to meet them without bias.
Developing the Agile Mind: Interventions and Neuroplasticity#
The observation that some leaders thrive in extreme complexity while others buckle under pressure highlights distinct individual variances in baseline cognitive flexibility. While demographic factors and prior experience play roles, the underlying neurobiological capacity for set-shifting is not static; it is highly subject to neuroplasticity. The human brain can be structurally and functionally modified to resist stress-induced PFC decoupling and maintain top-down executive control. This training generally targets two distinct vectors: modifying the physiological threshold of the stress response, and explicitly strengthening the connectivity and efficiency of the top-down regulatory networks.
Stress Inoculation Training (SIT) and Structural Adaptation#
The most direct and biologically potent method to prevent the catecholaminergic hijacking of the prefrontal cortex is to elevate the threshold at which a stimulus is perceived as an uncontrollable threat. This is systematically achieved through Stress Inoculation Training (SIT).
SIT operates on the biological principle of hormesis, the phenomenon whereby a beneficial, strengthening effect results from exposure to low doses of an otherwise toxic agent. Events that induce very high levels of uncontrolled, toxic stress can overwhelm coping capacities, leading to trauma and context rigidity. Conversely, events that induce no stress provide zero resilience-building value. However, exposure to moderately stressful events (tolerable stress) in a highly controlled environment allows the brain to rehearse coping mechanisms, effectively “inoculating” the individual against future, more severe trauma by altering how they appraise stress.
Neuroimaging studies reveal that coping with early or controlled life stress triggers profound developmental cascades that result in enduring architectural changes in the brain. Specifically, stress inoculation expands the surface area of the ventromedial prefrontal cortex (vmPFC). It increases white matter myelination, inferred from diffusion tensor magnetic resonance imaging, in the critical neural pathways connecting the PFC to the amygdala. The vmPFC is a region that broadly regulates arousal, supports emotional learning, and contributes to physiological resilience.
By thickening and myelinating these top-down inhibitory pathways, SIT ensures that the prefrontal cortex can send faster, significantly more robust signals to dampen amygdala hyperreactivity during an acute crisis. The leader’s brain is fundamentally, physically rewired. When a massive prediction error occurs in the real world, the reinforced vmPFC prevents the D1 receptor overload in the dlPFC, allowing the leader to maintain goal-directed cognition rather than defaulting to habitual panic. This is precisely why elite military units, commercial aviators, and specialized crisis communication teams rely so heavily on high-fidelity simulation training; they are not just learning procedures, they are literally myelinating their cognitive flexibility networks to ensure rapid decision-making under extreme duress.
Mindfulness, Non-Reactivity, and the Executive Buffer#
While SIT directly addresses the physiological stress threshold, distinct cognitive training paradigms aim to directly enhance the computational efficiency of the L-FPN and M-CIN networks. In recent years, an expansive, highly rigorous body of research has demonstrated the profound efficacy of Mindfulness-Based Stress Reduction (MBSR) and related cognitive therapies in bolstering cognitive flexibility.
Historically, mindfulness was viewed through an abstract or purely philosophical lens, but computational psychiatry and functional neuroimaging have elucidated its precise operational mechanisms. Regular mindfulness training reduces baseline activity in the amygdala and demonstrably strengthens functional connectivity in cortical areas responsible for emotional control and executive function. Crucially, the specific, measurable mechanism through which mindfulness improves cognitive flexibility is defined as “non-reactivity”. Path analysis of individuals undergoing MBSR training shows that treatment-induced changes in cognitive flexibility at post-treatment are fully mediated by non-reactivity scores measured midway through the intervention.
Non-reactivity is the trained cognitive capacity to observe internal thoughts, emotional states, and external sensory inputs without immediate judgment, reflexive engagement, or automatic response execution. In the context of predictive coding and the Bayesian brain, non-reactivity acts as a vital epistemic buffer. When a massive prediction error occurs, the standard, stressed brain rushes to active inference, immediately reacting to reduce the painful psychological discomfort of uncertainty. Non-reactivity allows the leader to tolerate the prediction error and the associated uncertainty, suspending the habitual, model-free response just long enough for the dlPFC and dACC to evaluate the incoming data, shift mental sets, and generate a novel, goal-directed strategy.
By rigorously practicing non-reactivity, individuals train the Salience Network to regulate its switching functions more efficiently, ensuring that conflict and uncertainty do not automatically trigger cognitive rigidity and threat priors. This trained ability to separate the stimulus from the response is the foundational neural pillar of adaptive leadership in chaotic environments.
The Corporate Imperative: Threat-Rigidity vs. Threat-Flexibility#
The neurobiology of the individual leader inextricably dictates the resilience and agility of the macro-organization. Consequently, the modern corporate environment increasingly views cognitive flexibility not as a peripheral “soft skill,” but as a core, non-negotiable requirement for strategic survival. Reports spanning from 2024 to 2026, including the World Economic Forum’s Future of Jobs Report, rank cognitive flexibility among the most critical skills for professional success in the 21st century.
When crises affect organizational structures, a paradoxical tension emerges between threat-rigidity and threat-flexibility. On one hand, the crisis induces profound emotional pressure. It severely constrains cognitive resources, pushing the workforce into risk-avoidance, inactivity, and strict adherence to familiar routines, the organizational equivalent of habit-based processing. On the other hand, a crisis presents an unmissable opportunity for innovation, proactive change, and the dismantling of obsolete paradigms to find safer outcomes.
Extensive research, including field surveys and scenario-based experiments in companies undergoing crises, indicates that the determining variable between organizational paralysis and a successful strategic pivot is the cognitive flexibility of its leadership apparatus. Chief Executive Officers possessing high cognitive flexibility are uniquely capable of driving “organizational ambidexterity”, the critical capability to simultaneously exploit current competitive advantages while rapidly exploring and developing innovations to meet shifting environments. Leaders capable of advanced perspective shifting and adaptive thinking act as regulatory neural nodes for their entire teams.
By consistently demonstrating non-reactivity, implementing clear heuristics (as evidenced in the Apollo 13 crisis), and communicating with total transparency, effective crisis leaders mitigate the transmission of toxic stress downward through the corporate hierarchy. This preserves the collective prefrontal capacity of the workforce, fostering an environment where employees feel psychologically safe enough to engage in proactive, goal-directed behavior despite the surrounding ambiguity.
Furthermore, recent expert consensus emphasizes that targeted cognitive flexibility training, incorporating task-switching protocols and complex set-shifting paradigms such as modified Stroop tasks, has strong potential to ameliorate executive function deficits and improve adaptive behavior in real-world settings. The Stroop effect, for instance, helps researchers understand how individuals allocate attention and manage cognitive resources when processing conflicting information, serving as both an assessment and a training tool for executive control. When organizations invest heavily in simulation-based learning, reflexivity loops, and personalized, adaptive cognitive training, they are not merely teaching theoretical crisis management; they are explicitly fortifying the physical neural architecture required to navigate the next global disruption.
Synthesis and Strategic Outlook#
The architecture of crisis leadership is inextricably bound to the neurochemical, structural, and network-level dynamics of the human brain. When global disruptions hit, the natural, evolutionary default is a catecholamine-driven retreat from the sophisticated prefrontal cortex into the rigid, habitual processing centers of the striatum and amygdala. In operational environments characterized by volatile prediction errors, high stakes, and profound uncertainty, this biological default reliably leads to catastrophic leadership failure, manifesting behaviorally as either the micromanagement of obsolete models or paralyzing, fear-driven inaction.
However, a deep examination of the neuro-mechanics of leadership reveals that this trajectory is decidedly not inevitable. Through the sophisticated, synchronized orchestration of the Salience Network, the Central Executive Network, and the Default Mode Network, the agile mind executes rapid set-shifting, ruthlessly abandoning failing heuristics and generating novel, goal-directed frameworks in real-time. The Bayesian brain can be explicitly conditioned to interpret environmental uncertainty not as a rigid threat but as a critical signal for necessary belief updating and epistemic foraging.
Master architects of leadership, as evidenced by paradigms such as Flight 1549 and Apollo 13, employ mechanisms ranging from the intentional millisecond pause to tight boundary heuristics. They bypass the neurochemical hijack through Naturalistic Decision-Making and the operational sequencing of the OODA loop. More importantly, this cognitive agility is a highly trainable biological asset. Through Stress Inoculation Training, which physically myelinates inhibitory neural pathways, and mindfulness interventions, which cultivate executive non-reactivity, leaders can fundamentally and permanently alter their neurobiological baseline.
In doing so, they forge an agile mind capable of thriving in the eye of the storm, transforming the chaos, uncertainty, and prediction errors of crises into structured avenues for unprecedented organizational resilience, adaptive innovation, and enduring success.
References#
- Shields, G. S., Sazma, M. A., & Yonelinas, A. P. (2016). The effects of acute stress on core executive functions: A meta-analysis and comparison with cortisol. Neuroscience and biobehavioral reviews, 68, 651-668. https://doi.org/10.1016/j.neubiorev.2016.06.038
- Dang, J. (2017). Commentary: The effects of acute stress on core executive functions: A meta-analysis and comparison with cortisol. Frontiers in Psychology, 8, 1711.
- Arnsten A. F. (2015). Stress weakens prefrontal networks: molecular insults to higher cognition. Nature Neuroscience, 18(10), 1376-1385. https://doi.org/10.1038/nn.4087
- Arnsten A. F. (2015). Stress weakens prefrontal networks: molecular insults to higher cognition. Nature Neuroscience, 18(10), 1376-1385. https://doi.org/10.1038/nn.4087
- Hains, A. B., Yabe, Y., & Arnsten, A. F. (2015). Chronic Stimulation of Alpha-2A-Adrenoceptors With Guanfacine Protects Rodent Prefrontal Cortex Dendritic Spines and Cognition From the Effects of Chronic Stress. Neurobiology of stress, 2, 1-9. https://doi.org/10.1016/j.ynstr.2015.01.001
- Arnsten, A. F., Raskind, M. A., Taylor, F. B., & Connor, D. F. (2015). The Effects of Stress Exposure on Prefrontal Cortex: Translating Basic Research into Successful Treatments for Post-Traumatic Stress Disorder. Neurobiology of stress, 1, 89-99. https://doi.org/10.1016/j.ynstr.2014.10.002
- Datta, D., & Arnsten, A. F. Loss of Prefrontal Cortical Higher Cognition with Uncontrollable Stress: Molecular Mechanisms, Changes with Age, and Relevance to Treatment. Brain Sciences, 9(5), 113. https://doi.org/10.3390/brainsci9050113
- Arnsten AF. Stress signalling pathways that impair prefrontal cortex structure and function. Nature reviews. Neuroscience. 2009 Jun;10(6):410-422. DOI: 10.1038/nrn2648. PMID: 19455173; PMCID: PMC2907136.
- Goodman, J., Leong, K. C., & Packard, M. G. (2015). Glucocorticoid enhancement of dorsolateral striatum-dependent habit memory requires concurrent noradrenergic activity. Neuroscience, 311, 1-8. https://doi.org/10.1016/j.neuroscience.2015.10.014
- Gadberry, T. M., Goodman, J., & Packard, M. G. (2022). Chronic corticosterone administration in adolescence enhances dorsolateral striatum-dependent learning in adulthood. Frontiers in Behavioral Neuroscience, 16, 970304. https://doi.org/10.3389/fnbeh.2022.970304
- Goode, T. D., Leong, K. C., Goodman, J., Maren, S., & Packard, M. G. (2016). Enhancement of striatum-dependent memory by conditioned fear is mediated by beta-adrenergic receptors in the basolateral amygdala. Neurobiology of Stress, 3, 74. https://doi.org/10.1016/j.ynstr.2016.02.004
- Goodman, J., Ressler, R.L., & Packard, M.G. (2016). The dorsolateral striatum selectively mediates extinction of habit memory. Neurobiology of learning and memory, 136, 54-62.
- Roozendaal B, Okuda S, Van der Zee EA, McGaugh JL. Glucocorticoid enhancement of memory requires arousal-induced noradrenergic activation in the basolateral amygdala. Proceedings of the National Academy of Sciences of the United States of America. 2006 Apr;103(17):6741-6746. DOI: 10.1073/pnas.0601874103. PMID: 16611726; PMCID: PMC1458951.
- Santiago, M., Machado, A., & Cano, J. (1993). Regulation of the prefrontal cortical dopamine release by GABAA and GABAB receptor agonists and antagonists. Brain research, 630(1-2), 28-31. https://doi.org/10.1016/0006-8993(93)90638-4
- Floresco, S. B. (2013). Prefrontal dopamine and behavioral flexibility: Shifting from an “inverted-U” toward a family of functions. Frontiers in Neuroscience, 7, 46724. https://doi.org/10.3389/fnins.2013.00062
- Armbruster-Genc, Diana & Ueltzhöffer, Kai & Basten, Ulrike & Fiebach, Christian. (2012). Prefrontal Cortical Mechanisms Underlying Individual Differences in Cognitive Flexibility and Stability. Journal of Cognitive Neuroscience. 24. 2385-2399. 10.1162/jocn_a_00286.
- Jenni, N. L., Larkin, J. D., & Floresco, S. B. (2017). Prefrontal Dopamine D1 and D2 Receptors Regulate Dissociable Aspects of Decision Making via Distinct Ventral Striatal and Amygdalar Circuits. The Journal of neuroscience: the official journal of the Society for Neuroscience, 37(26), 6200-6213. https://doi.org/10.1523/JNEUROSCI.0030-17.2017
- Musaelyan, K., Yildizoglu, S., Bozeman, J., Du Preez, A., Egeland, M., Zunszain, P. A., Pariante, C. M., Fernandes, C., & Thuret, S. (2020). Chronic stress induces significant gene expression changes in the prefrontal cortex alongside alterations in adult hippocampal neurogenesis. Brain communications, 2(2), fcaa153. https://doi.org/10.1093/braincomms/fcaa153
- Moench, K. M., Breach, M. R., & Wellman, C. L. (2020). Prior stress followed by a novel stress challenge results in sex-specific deficits in behavioral flexibility and changes in gene expression in rat medial prefrontal cortex. Hormones and behavior, 117, 104615. https://doi.org/10.1016/j.yhbeh.2019.104615
- Sterrenburg, L., Gaszner, B., Boerrigter, J., Santbergen, L., Bramini, M., Elliott, E., Chen, A., M. Peeters, W. M., Roubos, E. W., & Kozicz, T. (2011). Chronic Stress Induces Sex-Specific Alterations in Methylation and Expression of Corticotropin-Releasing Factor Gene in the Rat. PLOS ONE, 6(11), e28128. https://doi.org/10.1371/journal.pone.0028128
- Zhang, Ying-Dan & Shi, Dong-Dong & Zhang, Sen & Wang, Zhen. (2023). Sex-specific transcriptional signatures in the medial prefrontal cortex underlying sexually dimorphic behavioural responses to stress in rats. Journal of Psychiatry and Neuroscience. 48. E61-E73. 10.1503/jpn.220147.
- Chand, G. B., & Dhamala, M. (2016). Interactions Among the Brain Default-Mode, Salience, and Central-Executive Networks During Perceptual Decision-Making of Moving Dots. Brain connectivity, 6(3), 249-254. https://doi.org/10.1089/brain.2015.0379
- Chand, G. B., Wu, J., Hajjar, I., & Qiu, D. (2017). Interactions of the Salience Network and Its Subsystems with the Default-Mode and the Central-Executive Networks in Normal Aging and Mild Cognitive Impairment. Brain connectivity, 7(7), 401-412. https://doi.org/10.1089/brain.2017.0509
- Chen, H., Li, Y., Liu, Q., Shi, Q., Wang, J., Shen, H., Chen, X., Ma, J., Ai, L., & Zhang, Y. M. (2019). Abnormal Interactions of the Salience Network, Central Executive Network, and Default-Mode Network in Patients With Different Cognitive Impairment Loads Caused by Leukoaraiosis. Frontiers in Neural Circuits, 13, 460986. https://doi.org/10.3389/fncir.2019.00042
- Seeley W. W. (2019). The Salience Network: A Neural System for Perceiving and Responding to Homeostatic Demands. The Journal of neuroscience: the official journal of the Society for Neuroscience, 39(50), 9878-9882. https://doi.org/10.1523/JNEUROSCI.1138-17.2019
- Menon V. (2023). 20 years of the default mode network: A review and synthesis. Neuron, 111(16), 2469-2487. https://doi.org/10.1016/j.neuron.2023.04.023
- Kemmerer D. (2025). Does the Default Mode Network Mediate an Ongoing Internal Narrative? An Evaluation of Menon’s (2023) Hypothesis. Journal of cognitive neuroscience, 37(12), 2676-2683. https://doi.org/10.1162/JOCN.a.66
- Azarias, F. R., Almeida, G. H., De Melo, L. F., Rici, R. E., & Maria, D. A. The Journey of the Default Mode Network: Development, Function, and Impact on Mental Health. Biology, 14(4), 395. https://doi.org/10.3390/biology14040395
- Marek, S., & Dosenbach, N. U. F. (2018). The frontoparietal network: function, electrophysiology, and importance of individual precision mapping. Dialogues in clinical neuroscience, 20(2), 133-140.
https://doi.org/10.31887/DCNS.2018.20.2/smarek
- Song, L., Wu, G., Zhang, J., Liu, B., Chen, X., Wang, J., Gu, X., Tian, B., Li, Y., Zhang, A., Ma, X., & Jiang, L. (2025). The changes in brain network functional gradients and dynamic functional connectivity in SeLECTS patients revealing disruptive and compensatory mechanisms in brain networks. Frontiers in Psychiatry, 16, 1584071. https://doi.org/10.3389/fpsyt.2025.1584071
- Sassenberg, Tyler & Safron, Adam & Deyoung, Colin. (2024). Stable Individual Differences from Dynamic Patterns of Function: Brain Network Flexibility Predicts Openness/Intellect and Intelligence. 10.1101/2024.01.05.574386.
- Caldinelli, C., & Cusack, R. (2021). The fronto‐parietal network is not a flexible hub during naturalistic cognition. Human Brain Mapping, 43(2), 750. https://doi.org/10.1002/hbm.25684
- Uddin, L. Q. (2021). Cognitive and behavioural flexibility: Neural mechanisms and clinical considerations. Nature Reviews Neuroscience, 22(3), 167-179. https://doi.org/10.1038/s41583-021-00428-w
- Borghesi F, Cipresso P. Decades of research on Cognitive, Affective, Behavioral, and Psychological Flexibility: A scientometric analysis of trends and knowledge clusters towards a shared definition of Mental Flexibility Heliyon, 2026; 12.
- Parr, Thomas & Friston, Karl. (2017). Uncertainty, epistemics and active inference. Journal of The Royal Society Interface. 14. 20170376. 10.1098/rsif.2017.0376.
- Lehmann, K., Bolis, D., Friston, K. J., Schilbach, L., D Ramstead, M. J., & Kanske, P. (2023). An Active-Inference Approach to Second-Person Neuroscience. Perspectives on Psychological Science, 19(6), 931. https://doi.org/10.1177/17456916231188000
- Kiverstein, J., & Miller, M. (2015). The embodied brain: towards a radical embodied cognitive neuroscience. Frontiers in human neuroscience, 9, 237. https://doi.org/10.3389/fnhum.2015.00237
- Monosov, I. E. (2020). How outcome uncertainty mediates attention, learning, and decision-making. Trends in Neurosciences, 43(10), 795. https://doi.org/10.1016/j.tins.2020.06.009
- Blankenstein, N. E. (2019). Neural tracking of subjective value under riskand ambiguity in adolescence. Cognitive, Affective & Behavioral Neuroscience, 19(6), 1364. https://doi.org/10.3758/s13415-019-00749-5
- Levy, Ifat & Snell, Jason & Nelson, Amy & Rustichini, Aldo & Glimcher, Paul. (2010). Neural Representation of Subjective Value Under Risk and Ambiguity. Journal of Neurophysiology. 103. 1036-47. 10.1152/jn.00853.2009.
- Felsenheimer, A., Baxter, T., Sangimino, M., & Park, S. (2025). The Role of Interoception in Felt Presence and Psychosis Risk. Psychopathology, 1-13. Advance online publication. https://doi.org/10.1159/000549423
- Snell, Lucy & Reynolds, Steven & Garner, Matthew & Pfeifer, Gaby & Morriss, Jayne. (2025). Exploring The Role of Interoception in Anxious Traits and Symptoms. 10.31234/osf.io/5f7q9_v1.
- Klein, Gary. (2001). Sources of Power: How People Make Decisions. 10.1061/(ASCE)1532-6748(2001)1:1(21).
- Mokline, Bechir & Ben Abdallah, Mohamed. (2021). Organizational resilience as response to a crisis: case of COVID-19 crisis. Continuity & Resilience Review. ahead-of-print. 10.1108/CRR-03-2021-0008.
- Muir, T., Poudyal, C. S., De Lima, R., & Otaki, F. (2025). Investigating organizational resilience in a medicine and health sciences university in United Arab Emirates. PloS one, 20(12), e0338728. https://doi.org/10.1371/journal.pone.0338728
- Zhang, S., Tian, Y., Liu, Q., & Wu, H. (2025). The neural correlates of novelty and variability in human decision-making under an active inference framework. eLife, 13, RP92892. https://doi.org/10.7554/eLife.92892
- Crowley, B. (2021). The OODA Loop. The Decision Lab. Retrieved June 3, 2026, from https://thedecisionlab.com/reference-guide/computer-science/the-ooda-loop
- Niklasson, L. & Riveiro, Maria & Johansson, Fredrik & Dahlbom, Anders & Falkman, Göran & Ziemke, Tom & Brax, Christoffer & Kronhamn, T. & Smedberg, M. & Warston, Håkan & Gustavsson, Per. (2008). Extending the scope of situation analysis. Information Fusion - INFFUS. 1 - 8. 10.1109/ICIF.2008.4632246.
- Mehta, R. K., & Parasuraman, R. (2013). Neuroergonomics: a review of applications to physical and cognitive work. Frontiers in human neuroscience, 7, 889. https://doi.org/10.3389/fnhum.2013.00889
- Dehais, Frédéric & Callan, Daniel. (2019). A Neuroergonomics Approach to Human Performance in Aviation. 10.4324/9780429492181-6.
- Bogataia, Olga. (2025). Cognitive flexibility and neuropsychological mechanisms of managerial decision-making under vuca conditions. Теоретичні і прикладні проблеми психології та соціальної роботи. 1. 332. 10.33216/2219-2654-2025-332-343-2-67.
- Tuna, Özlem. (2025). The Relationship between the VUCA Environment and Managers’ Decision-Making Styles. İş ve İnsan Dergisi. 12. 70-85. 10.18394/iid.1552841.
- Shih, P. C., Pérez-Santiago, Á., Peña, D., Wazne, D., & Román, S. (2025). Jumping to Conclusions: Mechanisms of Cognitive Control in Decision-Making Under Uncertainty. Behavioral Sciences (Basel, Switzerland), 15(2), 226. https://doi.org/10.3390/bs15020226
- Crum, A. J., Akinola, M., Martin, A., & Fath, S. (2017). The role of stress mindset in shaping cognitive, emotional, and physiological responses to challenging and threatening stress. Anxiety, stress, and coping, 30(4), 379-395. https://doi.org/10.1080/10615806.2016.1275585
- Keech, J.J., Hamilton, K. (2020). Stress Mindset. In: Gellman, M.D. (eds) Encyclopedia of Behavioral Medicine. Springer, Cham. https://doi.org/10.1007/978-3-030-39903-0_102001
- Lindsay, E. K., & Creswell, J. D. (2017). Mechanisms of mindfulness training: Monitor and Acceptance Theory (MAT). Clinical psychology review, 51, 48-59. https://doi.org/10.1016/j.cpr.2016.10.011
- Lindsay, Emily & Creswell, J. (2018). Mindfulness, acceptance, and emotion regulation: Perspectives from Monitor and Acceptance Theory (MAT). Current Opinion in Psychology. 28. 10.1016/j.copsyc.2018.12.004.
- Creswell, J. & Lindsay, Emily. (2014). How Does Mindfulness Training Affect Health? A Mindfulness Stress Buffering Account. Current Directions in Psychological Science. 23. 401-407. 10.1177/0963721414547415.
- Lindsay, E. K., Young, S., Smyth, J. M., Brown, K. W., & Creswell, J. D. (2018). Acceptance lowers stress reactivity: Dismantling mindfulness training in a randomized controlled trial. Psychoneuroendocrinology, 87, 63-73. https://doi.org/10.1016/j.psyneuen.2017.09.015
- Chin, Brian & Slutsky, Jerry & Raye, Julianna & Creswell, John. (2019). Mindfulness Training Reduces Stress At Work: A Randomized Controlled Trial. Mindfulness. 10. 1-12. 10.1007/s12671-018-1022-0.
- Creswell, J. D., Irwin, M. R., Burklund, L. J., Lieberman, M. D., Arevalo, J. M., Ma, J., Breen, E. C., & Cole, S. W. (2012). Mindfulness-Based Stress Reduction training reduces loneliness and pro-inflammatory gene expression in older adults: a small randomized controlled trial. Brain, behavior, and immunity, 26(7), 1095-1101. https://doi.org/10.1016/j.bbi.2012.07.006
- Tang, Y. Y., Hölzel, B. K., & Posner, M. I. (2015). The neuroscience of mindfulness meditation. Nature Reviews. Neuroscience, 16(4), 213-225. https://doi.org/10.1038/nrn3916
- Stanley, Elizabeth. (2014). Mindfulness‐Based Mind Fitness Training: An Approach for Enhancing Performance and Building Resilience in High‐Stress Contexts. 10.1002/9781118294895.ch50.
- Galante, J., Dufour, G., Vainre, M., Wagner, A. P., Stochl, J., Benton, A., Lathia, N., Howarth, E., & Jones, P. B. (2018). A mindfulness-based intervention to increase resilience to stress in university students (the Mindful Student Study): a pragmatic randomised controlled trial. The Lancet. Public health, 3(2), e72-e81. https://doi.org/10.1016/S2468-2667(17)30231-1
- Yuan, M., & Hu, Z. (2025). Enhancing academic resilience through mindfulness training: An experimental study with Chinese undergraduates and the mediating role of psychological flexibility. Frontiers in Psychology, 16, 1692295. https://doi.org/10.3389/fpsyg.2025.1692295
- Diamond, A., & Ling, D. S. (2019). Aerobic-Exercise and resistance-training interventions have been among the least effective ways to improve executive functions of any method tried thus far. Developmental cognitive neuroscience, 37, 100572. https://doi.org/10.1016/j.dcn.2018.05.001
- Diamond, Adele & Ling, Daphne. (2020). Review of the Evidence on, and Fundamental Questions About, Efforts to Improve Executive Functions, Including Working Memory. 10.1093/oso/9780199974467.003.0008.
- Lenze, E. J., Voegtle, M., Miller, J. P., Ances, B. M., Balota, D. A., Barch, D., Depp, C. A., Diniz, B. S., Eyler, L. T., Foster, E. R., Gettinger, T. R., Head, D., Hershey, T., Klein, S., Nichols, J. F., Nicol, G. E., Nishino, T., Patterson, B. W., Rodebaugh, T. L., Schweiger, J., … Wetherell, J. L. (2022). Effects of Mindfulness Training and Exercise on Cognitive Function in Older Adults: A Randomized Clinical Trial. JAMA, 328(22), 2218-2229. https://doi.org/10.1001/jama.2022.21680
- Hindin SB, Zelinski EM. Extended practice and aerobic exercise interventions benefit untrained cognitive outcomes in older adults: a meta-analysis. Journal of the American Geriatrics Society 2012; 60(1): 136-141.
- Fernandez, V. Behavioral Rigidity vs. Strategic Flexibility: Family Firms in a Global Crisis. World, 7(5), 87. https://doi.org/10.3390/world7050087
- Sarkar, Soumodip & Osiyevskyy, Oleksiy. (2017). Organizational change and rigidity during crisis: A review of the paradox. European Management Journal. 36. 10.1016/j.emj.2017.03.007.
- Tarody, David. (2016). Organizational ambidexterity as a new research paradigm in strategic management. Vezetéstudomány / Budapest Management Review. 39-52. 10.14267/VEZTUD.2016.05.04.
- Tong, K., Fu, X., Hoo, N. P., Mun, L. K., Vassiliu, C., Langley, C., Sahakian, B. J., & Leong, V. (2024). The development of cognitive flexibility and its implications for mental health disorders. Psychological Medicine, 54(12), 3203. https://doi.org/10.1017/S0033291724001508
- Roux, Etienne & Beccaria, Gavin & McIlveen, Peter. (2024). The role of cognitive flexibility in job search behaviour: a research agenda. International Journal for Educational and Vocational Guidance. 25. 1347-1364. 10.1007/s10775-024-09669-4.
- Kiss, Andreea & Libaers, Dirk & Barr, Pamela & Wang, Tang & Zachary, Miles. (2020). CEO cognitive flexibility, information search, and organizational ambidexterity. Strategic Management Journal. 41. 2200-2233. 10.1002/smj.3192.
- Resendiz, S. M., Hernandez, M., Murphy, M., Casey, S., Chui, M. A., Burnside, E. S., & Sweeney, W. A. (2026). Psychological safety in interdisciplinary teams: How leadership behaviors empower teams. Frontiers in Psychology, 17, 1768461. https://doi.org/10.3389/fpsyg.2026.1768461
- Colgate, Orla & Colgate, Mark. (2025). The Neurobiology of Effective Leadership: Integrating Polyvagal Theory with the Coaching Leadership Style. Administrative Sciences. 15. 461. 10.3390/admsci15120461.


